47.Обмін піримідинових регуляції синтезу, розклад піримідинових. Порушення.
![]()
![]()
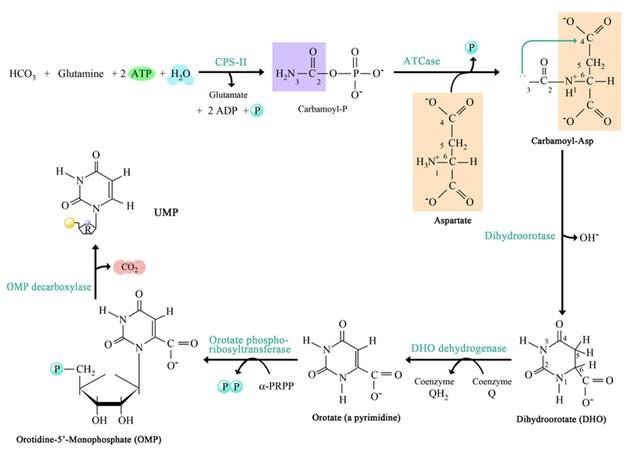
Это один из путей синтеза нуклеиновых кислот. В пиримидинах есть три молекулы нуклеотидов; это UTP, CTP и TTP. Путь синтеза пиримидина De novo можно объяснить следующими этапами.
1. Синтез карбамоилфосфата
2. Синтез карбамоил аспартата
3. Закрытие кольца для образования дигидрооротата
4. Окисление дигидро-оротата
5. Добавление рибозофосфатного фрагмента
6. Де Карбоксилирование с образованием UMP
![]()
Порушення синтезу піримідинових нуклеотидів. Спадкове порушення синтезу оротової кислоти, відоме як оротацидурія, характеризується утворенням її надлишку Кількість виведеної оротової кислоти в складі сечі при цьому може зростати до 1,0 – 1,5 г , що в 1 000 разів перевищує норму.
Оротатацидурія І типу- порушення піримідинового обміну, зумовлене дефіцитом двох ферментів:оротатфосфорибозилтрансферази, яка каталізує перетворення оротату на оротидилат, ідекарбоксилази, що каталізує перетворення оротидилату на уридинмонофосфат. Діти народжуються нормальними, але впродовж першого року життя в них розвивається мегалобластична анемія, згодом вони відстають у рості і розумовому розвитку. У сечі виявляють кристали оротової кислоти помаранчевого кольору (помаранчева кристалоурія).
Захворювання ставить дітей у залежність від зовнішніх джерел уридину. Вживання уридину чи цитидину призводить до зменшення утворення й екскреції оротової кислоти, відновлює нормальний ріст і розвиток. Таке лікування повинно продовжуватися впродовж усього життя.
Оратацидурія ІІ типурозвивається при нестачі тільки одного ферменту –оротидин-5-фосфатдекарбоксилази. У таких пацієнтів у сечі виявляють оротидин на відміну від І типу патології, при якій більшою мірою екскретується оротова кислота.
Вторинна оротова ацидурія(синдром Рейє) виникає при пошкодженні мітохондрій печінки. У результаті порушується утилізація карбамоїлфосфату, він включається в цикл синтезу оротової кислоти.
порушення катаболізму піримідинових нуклеотидів. Відомі порушення в робті двох ферментів цього метаболічного шляху. При недостатності піримідин-5’-нуклеотидази порушується відщеплення неорганічного фосфату від піримідинових мононуклеотидів і, відповідно, утворення нуклеозидів. Неактивна ізоформа зазначеного фермента виявлена в еритроцитах, там відбувається накопичення піримідинових НТФ, котрі інгібують пентозофосфатний шлях перетворення глюкози і створюють умови для гемолізу цих клітин крові.
Дигідропіримідиндегідрогеназа – фермент катаболізму піримідинів, який регулює його швидкість. Порушення роботи цього фермента супроводжується відхиленнями в функціонуванні нервової системи та діагностується на основі підвищення рівня вільних піримідинів – урацилу та тиміну в плазмі крові.
Регуляція синтезу піримідинових нуклеотидів.Контроль синтезу піримідинових нуклеотидів забезпечується на рівні двох регуляторних ферментів:
1. карбамоїлфосфатсинтетази, активність якої посилює ФРПФ, а алостеричними інгібіторами виступають УТФ – кінцевий продукт біосинтетичного процесу та пуринові нуклеотиди
2. аспартаткарбамоїлтрансферази,активатором якої слугує АТФ, а алостеричним інгібітором виступає ЦТФ.
Такий спосіб регуляції запобігає надлишковому синтезу не лише УМФ, а й усіх інших піримідинових нуклеотидів і забезпечує збалансоване утворення всіх чотирьох основних пуринових і піримідинових нуклеотидів, необхідних для синтезу РНК.